
【论著】| CDC20在肺腺癌组织中的表达及对肺腺癌细胞增殖和侵袭的影响研究
2024-06-27 中国癌症杂志 中国癌症杂志
CDC20在肺腺癌中高表达,CDC20高表达是肺腺癌患者不良预后的独立危险因素。CDC20能促进肺腺癌细胞增殖、迁移和侵袭。
[摘要] 背景与目的:肺腺癌具有早期发现难、肿瘤进展快及晚期手术切除率低等特点。尽管单药免疫治疗和免疫治疗联合化疗的相关研究在改善预后、克服耐药方面已初显成效,但是大部分肺腺癌患者从中获益仍有限。因此,迫切需要寻找具有相对较高灵敏度和特异度的新型生物标志物,以改善肺腺癌患者的预后。细胞分裂周期蛋白20(cell division cycle protein 20,CDC20)参与多种肿瘤的发生、发展,但在肺腺癌中的生物学作用及机制尚未明确。本研究旨在探究CDC20在肺腺癌中的表达情况及其对肺腺癌患者预后的预测价值,并分析CDC20对肺腺癌细胞增殖和侵袭能力的影响。方法:采用免疫组织化学(immunohistochemistry,IHC)检测CDC20在肺腺癌中的表达情况并结合生物信息学和临床病理学参数分析其与预后不良的相关性。采用Kaplan-Meier生存曲线描述CDC20对肺腺癌患者术后生存率的影响,采用COX多因素回归分析影响肺腺癌患者术后生存率的独立预后因素。通过受试者工作特征曲线分析CDC20表达在肺腺癌患者中的诊断价值。采用实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)和蛋白质印迹法(Western blot)检测人正常肺上皮细胞系BEAS-2B、人肺腺癌细胞系A549和H1299中CDC20的表达水平。细胞实验中,通过敲低肺腺癌细胞中的CDC20,分为si-NC(对照组)、si-CDC20#1(敲低组1)和si-CDC20#2(敲低组2)3个组。采用细胞计数试剂盒-8(cell counting kit-8,CCK-8)、克隆形成、transwell和划痕实验检测细胞增殖、迁移和侵袭能力。通过基因本体论(Gene Ontology,GO)功能和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路富集分析CDC20在肺腺癌中的生物学作用。通过基因集富集分析(gene set enrichment analysis,GSEA)CDC20在肺腺癌中可能的调控通路。本研究经河北省胸科医院伦理委员会批准(编号:2022051)。结果:生物信息学及IHC结果均显示,CDC20在肺腺癌组织中显著高表达(P<0.05)。生物信息学与临床参数分析结果均显示,CDC20高表达与患者预后不良相关。Kaplan-Meier生存分析和COX回归分析均显示,CDC20表达情况与患者术后生存率呈显著负相关(P<0.05)。敲低CDC20能抑制肺腺癌细胞增殖、迁移和侵袭(P<0.05)。GO功能、KEGG通路和GSEA结果均显示,CDC20与细胞周期相关。结论:CDC20在肺腺癌中高表达,CDC20高表达是肺腺癌患者不良预后的独立危险因素。CDC20能促进肺腺癌细胞增殖、迁移和侵袭。
[关键词] 细胞分裂周期蛋白20;肺腺癌;细胞增殖;细胞周期;细胞运动
[Abstract]Background and purpose: Lung adenocarcinoma has the characteristics of difficult early detection, rapid tumor progression and low surgical rep rate. Although studies on immunotherapy alone and immunotherapy combined with chemotherapy have shown initial success in improving prognosis and overcoming drug resistance, the majority of lung adenocarcinoma patients still receive limited benefits. Therefore, there is an urgent need to identify novel biomarkers with relatively high sensitivity and specificity to improve the prognosis of lung adenocarcinoma. Cell division cycle protein 20 (CDC20) is involved in the occurrence and development of various tumors, but its biological role and mechanism in lung adenocarcinoma remain unclear. The aim of this study was to investigate the expression of CDC20 in lung adenocarcinoma and its predictive value for the prognosis of patients with lung adenocarcinoma, and to further explore the effects of CDC20 on the proliferation and invasion capabilities of lung adenocarcinoma cells. Methods: Utilizing immunohistochemistry (IHC) to detect the expression of CDC20 in lung adenocarcinoma tissues, we analyzed its correlation with poor prognosis in combination with bioinformatics and clinicopathological parameters. Kaplan-Meier survival curves were employed to illustrate the impact of CDC20 on postoperative survival rates in lung adenocarcinoma patients. COX multivariate regression analysis was conducted to identify independent prognostic factors influencing postoperative survival rates. Additionally, receiver operating characteristic (ROC) curves were applied to assess the diagnostic value of CDC20 expression in lung adenocarcinoma patients. Real-time fluorescence quantitative polymerase chain reaction (RTFQ-PCR) and Western blot were used to measure the expression levels of CDC20 in normal human lung epithelial cells (BEAS-2B) and human lung adenocarcinoma cells (A549 and H1299). In cellular experiments, CDC20 was knocked down in lung adenocarcinoma cells, which were divided into three groups: si-NC (control group), si-CDC20#1 (knockdown group 1) and si-CDC20#2 (knockdown group 2). Cell counting kit-8 (CCK-8), colony formation, transwell and wound healing assays were conducted to assess cell proliferation, migration and invasion capabilities. Functional enrichment analysis using Geng Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways was conducted to explore the biological roles of CDC20 in lung adenocarcinoma. Finally, gene set enrichment analysis (GSEA) was employed to investigate potential regulatory pathways of CDC20 in lung adenocarcinoma. This study was approved by the Ethics Committee of Hebei Chest Hospital (Number: 2022051). Results: The results of both bioinformatics analysis and IHC demonstrated a significantly high expression of CDC20 in lung adenocarcinoma tissues (P<0.05). Both bioinformatics analysis and clinical parameter evaluation revealed a correlation between high CDC20 expression and poor patient prognosis. Kaplan-Meier survival analysis and COX regression analysis consistently indicated a significant negative correlation between CDC20 expression and postoperative survival rates in patients (P<0.05). Additionally, the expression levels of CDC20 were higher in human lung adenocarcinoma cell lines A549 and H1299 compared with BEAS-2B (P<0.05). Knockdown of CDC20 effectively inhibited the proliferation, migration and invasion of lung adenocarcinoma cells (P<0.05). The results of GO, KEGG pathways and GSEA consistently pointed to a relationship between CDC20 and cell cycle regulation. Conclusion: CDC20 is highly expressed in lung adenocarcinoma. High expression of CDC20 is an independent risk factor for poor prognosis of lung adenocarcinoma patient. CDC20 can promote the proliferation, migration and invasion of lung adenocarcinoma cells.
[Key words] Cell division cycle protein 20; Adenocarcinoma of the lung; Cell proliferation; Cell cycle; Cell movement
肺癌发病率和死亡率均较高,根据美国癌症协会的估计,到2023年底,可能有127 070人死于肺癌,占癌症总死亡率的20%[1-2]。针对肺腺癌的治疗方法已取得了长足进步,如免疫检查点抑制剂(immune checkpoint inhibitor,ICI)治疗和靶向治疗[3]。但由于肿瘤的异质性和复杂性,只有20%的局部晚期和转移性非小细胞肺癌患者可以从免疫治疗中获益[2]。因此,肺腺癌患者的预后仍然很差,5年生存率约为18%[4]。迫切需要寻找灵敏度和特异度更高的新型生物标志物,以改善肺腺癌患者的预后。
近年来,细胞分裂周期蛋白20(cell division cycle protein 20,CDC20)在多种肿瘤中的作用受到关注,因为其异常表达与恶性肿瘤的进展及不良预后密切相关,使其成为一种可能的肿瘤治疗靶点[5]。CDC20是调控有丝分裂后期启动和退出的关键有丝分裂因子,可间接影响细胞增殖[6]。抑制CDC20可增强替莫唑胺耐药胶质瘤细胞的敏感性,恢复上皮标志物E-钙黏蛋白(E-cadherin)的表达水平,降低间质标志物波形蛋白(vimentin)的表达[7]。在机制方面,之前的一项研究[8]指出,CDC20可与miRNA相互作用,从而对肝细胞癌产生影响,但是关于CDC20的生物信息学分析及临床结合基础研究尚未见报道。因此CDC20表达与肺腺癌的相关性及其在肺腺癌中的作用机制仍有待进一步探究。
本研究旨在通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)、高通量基因表达(Gene Expression Omnibus,GEO)数据库,应用生物信息学方法分析CDC20在肺腺癌中的表达及其与生存预后的相关性。通过基因集富集分析(gene set enrichment analysis,GSEA)挖掘CDC20可能参与的信号转导通路,揭示其在肺腺癌发生、发展中的作用及分子机制,最后通过组织、细胞实验来探索CDC20在肺腺癌中的潜在应用价值,从而探讨肺腺癌中CDC20的表达及临床预后意义,为肺腺癌的防治提供理论依据。
1 材料和方法
1.1 实验材料
从TCGA数据库中下载了具有临床数据的肺腺癌样本的每百万映射读取的每千碱基转录本的NA-seq片段数(FPKM),排除了没有临床数据的RNA-seq样本,然后将3级HTSeq-FPKM数据转换为每百万读取的转录本,共筛选出539例肺腺癌及58例癌旁正常组织样本;其中男性283例,女性256例;263例患者>65岁,276例患 者≤65岁。GEO数据集GSE115002、GSE31210、GSE68571、GSE31210和GSE116959用于验证肺腺癌及癌旁正常组织中mRNA的差异表达。另选取2015年1月—2020年12月于河北省胸科医院接受手术治疗并经病理学检查证实为肺腺癌的组织标本118例,另选取42例距离肿瘤5 cm以上的癌旁正常组织作为癌旁组,患者术前均未接受放化疗及其他免疫治疗。采用门诊复查、定期住院及电话回访等形式对118例患者进行随访,随访起止时间为2015年1月—2020年12月,共随访60个月,中位随访时间为48个月,无失访。本研究经河北省胸科医院伦理委员会批准(编号:2022051),患者均已签署知情同意书。人正常肺上皮细胞系BEAS-2B、人肺腺癌细胞系A549和H1299均购自中国医学科学院。实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)及反转录试剂盒均购自美国Sigma公司,PCR引物、CDC20的siRNA序列(siCDC20)及阴性对照序列(siNC)由上海吉玛制药技术有限公司合成,细胞计数试剂盒-8(cell counting kit-8,CCK-8)、二辛可宁酸(bicinchoninic acid,BCA)蛋白定量试剂盒均购自上海碧云天生物技术股份有限公司,TRIzol试剂和LipofectamineTM2000转染试剂均购自英国Abcam公司,TUNEL试剂盒购自武汉赛维尔生物科技有限公司,transwell小室购自美国NEST公司,兔抗人GAPDH一抗、兔抗人CDC20一抗及辣根过氧化物酶(horseradishperoxidase,HRP)标记的山羊抗兔免疫球蛋白G(immunoglobulin G,IgG)二抗均购自美国Sigma-Aldrich公司,SP试剂盒购自上海科欣生物技术研究所,DAB试剂和CDC20多克隆抗体均购自武汉三鹰生物技术有限公司。DMEM高糖细胞培养液、0.25%胰蛋白酶、RIPA裂解液和胎牛血清均购自美国Gibco公司。
1.2 方法
1.2.1 差异表达基因(differentially expressed genes,DEG)的筛选
基于TCGA数据库,使用R软件(3.6.2版)确定与CDC20共表达的基因。采用Spearman相关性分析探究CDC20与共表达基因的相关性,|r|>0.5和P<0.001表示CDC20与共表达基因显著相关。将CDC20(50%临界值)分为高表达和低表达组。使用R包DESeq2[1.38.1][9]确定两组之间的表达差异。DEG的阈值为P<0.05,|lgFC|>1,FC即变化倍数(fold change)。使用ggplot2 R包对DEG和共表达基因的数据进行可视化。
1.2.2 肺腺癌中CDC20表达与临床病理学特征及预后的相关性分析
采用Wilcoxon秩和检验(连续变量)或Pearson χ2检验比较CDC20高表达组和低表达组的临床病理学特征。通过受试者工作特征(receiver operator characteristic,ROC)曲线分析CDC20对肺腺癌诊断的预测价值。采用R包pROC[1.18.0]分析ROC曲线,并计算ROC曲线的曲线下面积(area under curve,AUC)。R包survival[3.3.1]用于生存资料的统计分析、比例风险假设检验并进行COX回归分析。使用R包rms构建nomogram相关模型列线图和校准图,并进行分析及可视化[10]。
1.2.3 富集分析
使用R包clusterProfiler[4.4.4][11]对DEG绘制基因本体论(Gene Ontology,GO)分析,包括生物过程(biological process,BP)、细胞成分(cellular component,CC)、分子功能(molecular function,MF)及京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG),P <0.05为差异有统计学意义。为评估CDC20高表达组和低表达组之间信号转导通路成分差异的重要性,使用R包clusterProfiler[3.8.0]对TCGA中的CDC20表达数据进行GSEA,以分子特征数据库(Molecular Signature Database,MsigDB)中的c2.cp.kegg. v7.0.symbols.gmt数据集作为功能基因集,P<0.05、错误发现率(false discovery rate,FDR)小于0.25和标准化富集评分(|NES|)>1被认为是显著富集。
1.2.4 免疫组织化学(immunohistochemistry,IHC)
组织石蜡切片、脱蜡、水化、抗原修复、渗透、封闭等操作后,给予一抗(CDC20抗体1∶200)过夜,给予生物素化通用二抗工作液,加入辣根过氧化物酶标记的链霉卵白素工作液DAB染色,苏木精复染,脱水封固。按染色强度:无色计0分,淡黄色计1分,棕黄色计2分,棕褐色计3分;按染色细胞百分比:<5%计为0分,5%~25%计1分,26%~50%计2分,>50%计3分。IHC结果:染色指数(SI)=染色强度×染色细胞。判定结果:0~1分为阴性,2~4分为弱阳性,5~7分为阳性,≥8分为强阳性。
1.2.5 细胞培养及转染
H1299、A549细胞和BEAS-2B细胞分别培养在含有10%胎牛血清、1%青-链霉素双抗的DMEM和RPMI-1640培养基中,于37 ℃、CO2体积分数为5%的培养箱中常规培养。当H1299和A549细胞融合度达到60%以上时,按脂质体LipofectamineTM2000说明书进行转染操作,并设置阴性对照组(si-NC)和转染靶向CDC20的siRNA组(siCDC20#1组和siCDC20#2组),用于后续研究。同时应用RTFQ-PCR与蛋白质印迹法(Western blot)技术检测各组A549和H1299细胞中CDC20的表达变化,以评估转染效率。
1.2.6 RTFQ-PCR
TRIzol法提取细胞总RNA,反转录试剂盒将RNA反转录为cDNA。采用SYBR RTFQ-PCR试剂盒进行PCR实验。以GAPDH作为内参,用2-ΔΔCt法计算CDC20 mRNA的相对表达量。CDC20的上游引物序列为5’-GGAGTGCAAGCTCTGGTGACATC-3’,下游引物序列为5 ’ -GGTGCCCACA GCCAAGTAGTTG-3’ ;GAPDH的上游引物序列为5’-AAGGAGTAAGA CCCCTGGACCA-3’,下游引物序列为5’-GCAAC TGTGAGCAGGGGAGATT-3’。
1.2.7 Western blot检测CDC20蛋白水平
各组细胞经预冷的磷酸缓冲盐溶液(phosphate-buffered saline,PBS)洗3次后,吸弃PBS。加入细胞裂解液在冰上裂解细胞以提取细胞中的总蛋白,通过BCA法测定蛋白浓度,经12%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecylsulphate polyacrylamide gel electrophoresis,SDS-PAGE)分离蛋白。电泳结束后将蛋白转移至PVDF膜上,再用脱脂奶粉封闭1 h,含有吐温-20三乙醇胺缓冲盐溶液(tris-buffered saline Tween,TBST)洗膜。将膜与GAPDH一抗(稀释比例:1∶1 000)、CDC20一抗(稀释比例:1∶1 000)4 ℃下温育过夜,将膜与HRP标记二抗(稀释比例:1∶2 000)常温温育1 h后,加入电化学发光(electrochemical luminescence,ECL)液曝光拍照。
1.2.8 细胞克隆形成实验检测细胞增殖能力
调整细胞密度至500个/孔接种于6孔板上,于37 ℃、CO2体积分数为5%的培养箱中培养3~4 d换液,培养10~14 d。4%多聚甲醛溶液固定15 min,结晶紫染色15 min,通过显微镜观察并拍照。
1.2.9 CCK-8检测细胞增殖能力
调整细胞密度至5×106个/mL,每孔100 μL接种于96孔板上,于37 ℃、CO2体积分数为5%的培养箱中培养24、48及72 h后,每孔加入10 μL CCK-8溶液继续温育2 h后,采用酶标仪测定450 nm波长处的吸光度(D)值。
1.2.10 划痕实验检测细胞迁移能力
计数并以2×105个/孔的细胞密度接种于6孔板上,待细胞融合度达到90%后,用移液器吸嘴于6孔板中划2条直线,PBS漂洗掉表面悬浮细胞,培养箱培养24 h后,计算划痕愈合率。
1.2.11 Transwell实验检测细胞侵袭能力
用不含胎牛血清的DMEM培养基重悬细胞,调整细胞密度至5×105个/mL,接种至含24孔板的transwell小室上室,将600 μL含10%胎牛血清的培养基加入下室,48 h后取出小室,甲醇溶液固定30 min,0.1%结晶紫染色,PBS洗净后在显微镜下拍照计数。
1.3 统计学处理
采用R Graphpad 9.0软件对数据进行统计分析和作图。计量资料以x±s表示,先进行方差齐性检验,临床预后分析采用Kaplan-Meier法。P <0.05为差异有统计学意义。数据采用Shapiro-Wilk法进行正态分布检验,符合正态分布的计量数据以x±s表示,两组间比较采用t检验,多组间比较采用单因素方差分析,计数资料以相对数描述,组间差异比较采用χ2检验,生存率计算用Kaplan-Meier法,并用log-rank检验比较生存率曲线。采用COX比例风险回归模型进行单因素和多因素分析。
2 结 果
2.1 数据库中CDC20表达的分析
通过TCGA数据库分析显示,CDC20 mRNA在多种肿瘤中呈显著高表达,其中肺腺癌组织中CDC20的表达明显高于癌旁组织(图1A),58对肿瘤配对样本中CDC20亦较正常表达更显著(P<0.001,图1B)。另外5个GEO数据集(GSE115002、GSE31210、GSE68571、GSE31210和GSE116959)进一步验证了CDC20在肺腺癌中高表达(图1C~1G)。
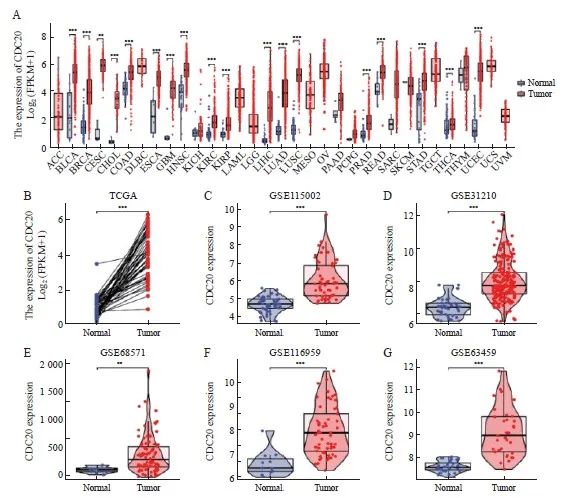
图1 CDC20在多种类型肿瘤中高表达
Fig. 1 CDC20 is highly expressed in multiple tumor types
A: Differential expression of CDC20 in pan-cancer samples from the TCGA database; B: CDC20 expression in lung adenocarcinoma and matched normal tissues in the TCGA databases; C-G: The CDC20 mRNA expression between lung adenocarcinoma and normal tissues based on data from GSE115002 (C), GSE31210 (D), GSE68571 (E),GSE116959(F) and GSE116959(G) dataset. **: P<0.01, compared with normal. ***: P<0.001, compared with normal.
2.2 TCGA数据库中CDC20的表达与临床病理学特征的相关性
通过TCGA数据库分析发现,CDC20表达的增加与临床分期(P<0.01,图2A)、T分期(P<0.001,图2B)、N分期(P<0.001,图 2C)、年龄(P<0.01,图2E)和性别(P<0.01,图2F)显著相关。
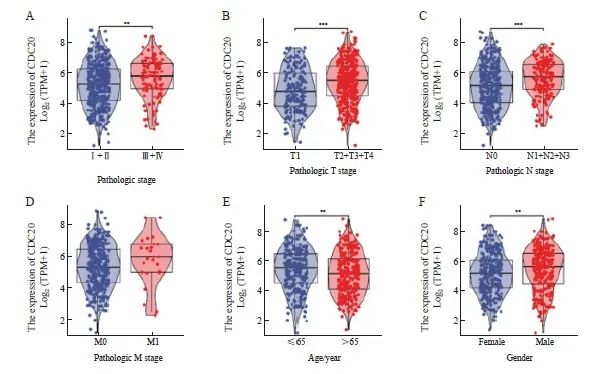
图2 CDC20的表达与临床病理学特征的相关性
Fig. 2 Association of CDC20 expression levels with clinicopathological features
A: Pathological stage; B: T stage; C: N stage; D: M stage; E: Age; F: Gender. ***: P<0.01; ***: P<0.001.
2.3 肺腺癌中CDC20的表达与临床病理学特征的相关性
CDC20高表达与临床分期、肿瘤大小、肿瘤分化程度和淋巴结转移有关(P<0.05,表1)。进一步分析发现,CDC20 mRNA高表达与更差的总生存期(overall survival,OS)和疾病特异性生存期(disease-specific survival,DSS)显著相关(图3A~3B)。采用ROC曲线评价CDC20 mRNA在肺腺癌中的诊断价值(图3C),AUC为0.979,提示CDC20 mRNA对肺腺癌具有较高的诊断效能。COX多因素回归分析结果显示,T分期、N分期和CDC20高表达被确定为独立的预后因素(图3D)。结合CDC20表达、临床分期来构建预测列线图模型,结果显示,列线图模型具有较高的准确度(C指数=0.693,图3E~3F)。
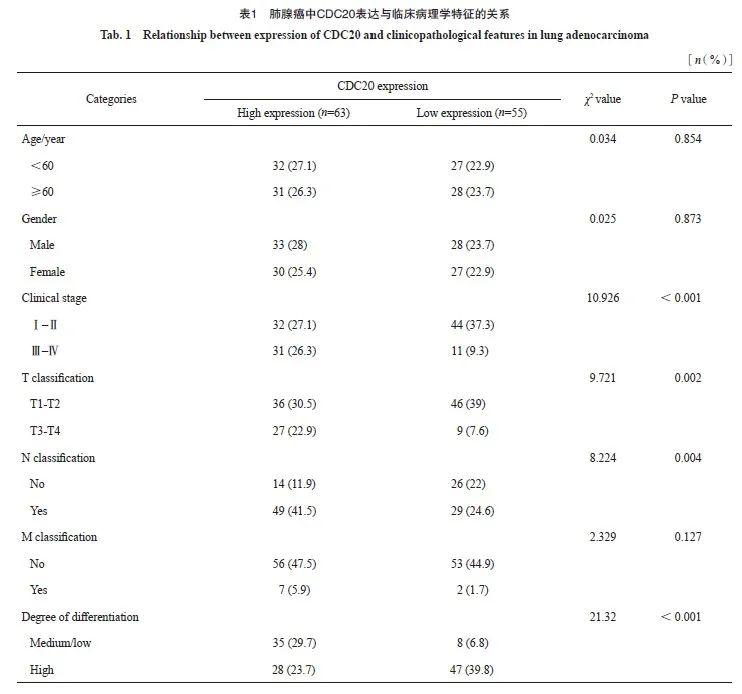
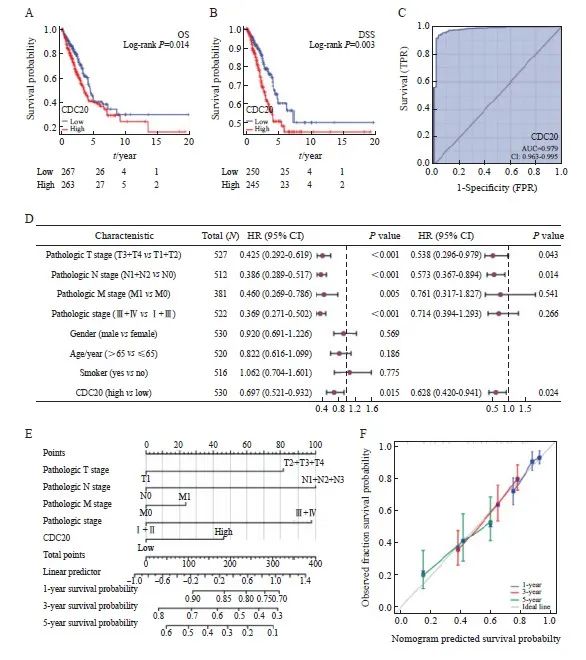
图3 CDC20表达在肺腺癌中的预后价值
Fig. 3 Prognostic value of CDC20 expression in lung adenocarcinoma
A, B: Survival curve; C: ROC diagnostic curve; D: Forest map of OS based on COX multivariate regression analysis; E: OS based nomograms and calibration curves; F: The calibration curve of nomogram.
2.4 肺腺癌中CDC20的表达与预后的相关性
通过分析118例肺腺癌组织和42例癌旁正常组织的IHC结果,发现CDC20表达定位于细胞质,多呈棕色或黄色(图4A),与癌旁正常组织相比,肺腺癌组织中CDC20的表达水平显著升高(P<0.001,图4B~4C)。肺腺癌患者的预后显示,CDC20表达与患者的OS率呈显著负相关(P <0.001,图4D)。COX多因素回归分析结果显示,CDC20高表达、临床分期、远处转移和淋巴结转移为独立的预后因素(表2)。
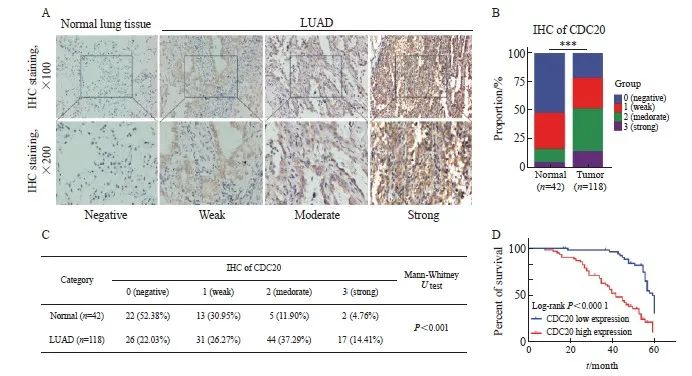
图4 肺腺癌中CDC20的表达与预后的相关性
Fig. 4 Correlation between CDC20 expression and prognosis in lung adenocarcinoma
A: IHC for detecting CDC20 expression in lung adenocarcinoma tissue and Normal lung tissue; B, C: Statistical analysis of tissue IHC scores; D: Kaplan-Meier analysis of CDC20 expression and OS based on lung adenocarcinoma tissue. ***: P<0.001, compared with normal.
2.5 CDC20在肺腺癌细胞中的表达及转染效果验证
通过RTFQ-PCR和Western blot检测CDC20 mRNA及蛋白在A549、H1299细胞系和人正常支气管上皮细胞系BEAS-2B中的表达水平。结果显示,与BEAS-2B细胞组相比,A549细胞和H1299细胞株中CDC20 mRNA及蛋白表达水平均显著增加(图5A,P <0.001);沉默CDC20基因表达后,与siNC组相比,siCDC20#1组和siCDC20#2组CDC20的表达水平均显著降低(图5B~5C,P <0.001),转染CDC20基因明显有效,可以进行后续细胞学功能试验。
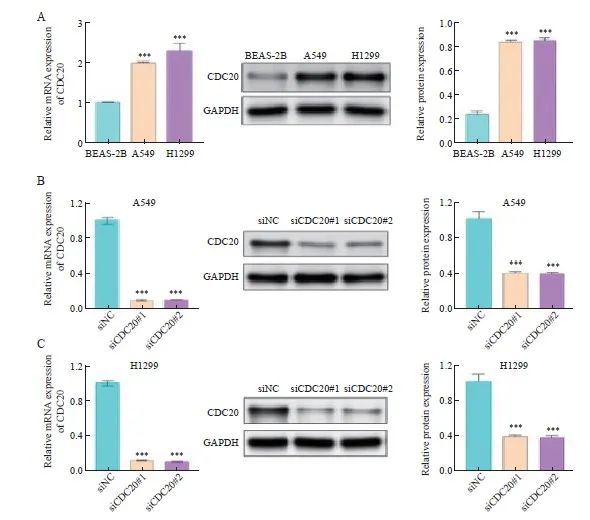
图5 肺腺癌细胞中CDC20 mRNA和蛋白的表达水平
Fig. 5 Expression levels of CDC20 mRNA and protein in lung adenocarcinoma cells in different groups
A: Expression levels of CDC20 mRNA and protein in BEAS-2B, A549 and H1299 cells; B: Relative expression levels of CDC20 mRNA and protein in A549 cells with CDC20 knockdown; C: Relative expression levels of CDC20 mRNA and protein in H1299 cells with CDC20 knockdown. ***: P <0.001, compared with BEAS-2B or siNC.
2.6 沉默CDC20对肺腺癌细胞增殖的影响
克隆形成实验显示,与siNC组相比,siCDC20#1组和siCDC20#2组细胞形成了更少的克隆数(图6A~6B)。CCK-8实验结果显示,与siNC组相比,siCDC20#1组和siCDC20#2组能够显著抑制A549和H1299细胞增殖(图6C~6D)。
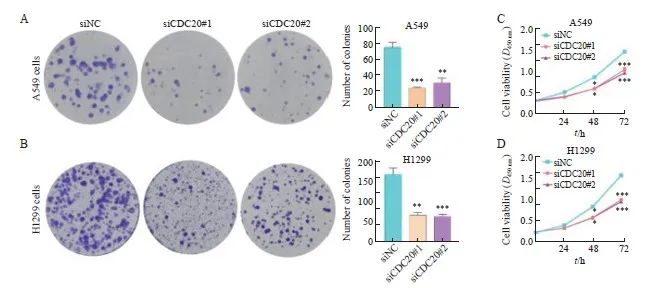
图6 沉默CDC20对A549和H1299细胞增殖能力的影响
Fig. 6 Effect of CDC20 silencing on the proliferation ability of A549 and H1299 cells
A, B: Colony formation assay showed that knockdown of CDC20 inhibited the proliferation of A549 and H1299 cells; C, D: CCK-8 assay showed that knockdown of CDC20 inhibited the proliferation of A549 and H1299 cells. *: P<0.05, compared with siNC; **: P<0.01, compared with siNC; ***: P<0.001, compared with siNC.
2.7 沉默CDC20对肺腺癌细胞迁移和侵袭的影响
划痕实验显示,与siNC组相比,siCDC20#1组和siCDC20#2组中A549和H1299细胞迁移能力明显降低(图7A~7B)。Transwell实验结果显示,与siNC组相比,siCDC20#1组和siCDC20#2组中A549和H1299细胞侵袭能力明显降低(图7C~7D)。
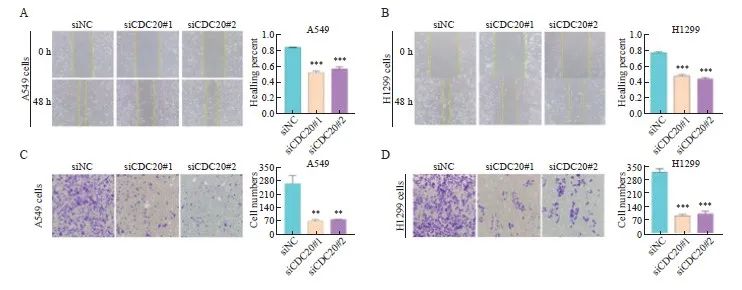
图7 沉默CDC20对A549和H1299细胞迁移和侵袭能力的影响
Fig. 7 Effect of CDC20 silencing on the migration and invasion of A549 and H1299 cells
A, B: Effect of CDC20 silencing on the migration ability of A549 and H1299 cells; C, D: Effect of CDC20 silencing on the invasion of A549 and H1299 cells. **:P<0.01, compared with siNC; ***: P<0.001, compared with siNC.
2.8 肺腺癌中CDC20相关基因功能分析
差异分析以火山图展示|log2(FC)|>1和p.adj<0.05阈值下的分子数目,火山图显示,满足条件的ID有9 593个,其中,高表达(log2FC为正)的数目有2 203个,低表达(log2FC为负)的数目有7 390个(图8A);共表达热图显示了与CDC20正负相关的前5个基因(图8B)。为研究与CDC20相关的生物学功能和信号通路,基于MSigDB Collection富集结果中的显著差异(调整后P<0.05,FDR<0.25),将CDC20高表达组和低表达组中的其他转录组基因进行了GSEA富集分析,发现与其相关的通路主要包括细胞周期、细胞周期有丝分裂、E2F、PLK1和FOXM1等通路(图8C)。KEGG信号通路结果显示,DEG主要富集在细胞周期、分泌胆汁过程、唾液分泌及蛋白质消化吸收等方面(图8D)。GO富集分析显示,DEG主要富集在纤毛运动、微管运动、细胞骨架运动活性及激素活动等方面(图8E)。
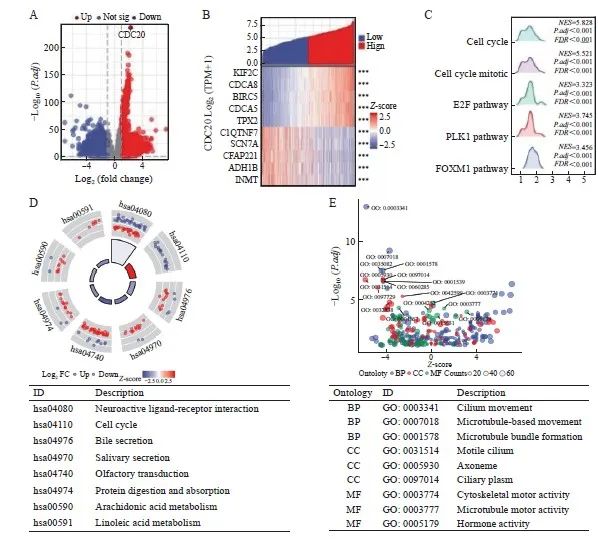
图8 DEG结果及功能富集分析
Fig. 8 Results of DEG and functional enrichment analysis
A: Volcano plot of DEG (red: upregulation; blue: downregulation); B: Heatmap of the correlation between CDC20 expression and the top 5 DEG; C: Results of GSEA enrichment analysis of DEG; D: KEGG pathway analysis; E: GO functional annotation analysis of biological process (BP), cellular component (CC) and molecular function (MF).
3 讨 论
肺腺癌存在恶性程度高、预后差及复发率高等特点[12-13]。为探索肺腺癌发生、发展的内在机制,开展了大量的基础和临床研究,对肺腺癌的治疗也有了较大进步,但全球肺腺癌的发病率和死亡率仍居高不下[14-15]。因此,急需寻找一种新的肺腺癌预后生物标志物,以指导个体化治疗和优化治疗策略。
CDC20是细胞周期相关蛋白之一,并作为晚期促进复合体的调节因子,在染色体分离和有丝分裂期间发挥关键的调控作用[16, 17]。已有研究显示,CDC20在胰腺癌[18]、乳腺癌[19]、前列腺癌[20]、结直肠癌[21]、肝细胞癌[22]、胶质瘤[23]、胃癌[24]、膀胱癌[25]及宫颈癌[26]等肿瘤中发挥作用,但其作用的具体分子机制,特别是与肺腺癌关系的相关研究报道少见。
有研究[27]表明,在恶性肿瘤中,CDC20作为主要的肿瘤启动子,可促进多种肿瘤的发生、发展,并可作为启动子调控G1/S转化的进展和癌细胞的存活。此外,Zheng等[28]的研究表明,CDC20除了启动外,对肿瘤的维持也至关重要,这些因素共同导致CDC20表达水平升高的患者预后不良。另有研究报道,CDC20在胰腺癌[18]、乳腺癌[19]、结直肠癌[21]、胶质母细胞瘤[23]及膀胱癌[25]等肿瘤中高表达并且与肿瘤的发生、发展密切相关。本研究首先对公共数据库进行挖掘,发现CDC20在肺腺癌中显著高表达,进一步通过实验验证发现CDC20在肺腺癌组织和细胞系中的表达均上调,与生物信息学预测结果保持一致,且CDC20高表达与临床分期、肿瘤大小和淋巴结转移有关,其中Kaplan-Meier生存分析显示,CDC20高表达的肺腺癌患者的死亡风险相对较高,COX单因素和多因素回归分析及列线图模型进一步证实CDC20是肺腺癌患者临床预后的独立预测因子,最终通过ROC曲线和校准曲线验证显示,CDC20建立的预后模型具有良好的效能。因此,CDC20除了作为判断预后的重要标志物外,还有可能成为肺腺癌的治疗靶点。
CDC20作为纺锤体组装检查点的靶点,在细胞周期调控中发挥重要作用。CDC20高表达使肿瘤细胞绕过纺锤体组装检查点的监视,提前退出有丝分裂过程,导致基因组不稳定[29],因此不难推测组织中表达过高的CDC20可能成为肿瘤发生、发展的危险因素。CDC20高表达还与Wnt/β-catenin信号通路激活相关,而Wnt/β-catenin信号通路与癌症进展密切相关[30-31]。为进一步了解CDC20的生物学功能和机制,我们在A549和H1299细胞中进行了相关体外实验。
本研究结果显示,敲减CDC20后A549和H1299细胞增殖、迁移和侵袭能力均显著降低,提示CDC20可能在调控肺腺癌细胞增殖、迁移和侵袭过程等方面发挥关键作用。但CDC20调控A549细胞和H1299细胞增殖、迁移和侵袭过程是否与Wnt/β-Catenin信号通路的调控有关,还需后续实验验证。
为进一步研究CDC20相关信号通路,我们通过GSEA分析TCGA数据库中CDC20表达水平不同的肺腺癌样本,发现CDC20与细胞周期、E2F靶点通路显著相关,而E2F是一个复杂的转录调节因子家族,在保护细胞免受细胞周期引起的基因组错误和异常增殖中发挥关键作用[32]。Zhang等[33]研究发现,E2F基因过表达与肺癌患者的不良预后相关。因此,CDC20可能通过参与E2F相关通路调控肺癌细胞的细胞周期,从而导致肺癌的发生、发展。但CDC20是否参与E2F相关通路调控肺癌细胞的细胞周期还需后续实验验证。综合上述研究结果及国内外相关报道,证实肿瘤的发生、发展是多方面因素决定的,因此对CDC20功能及通路研究将可能加深对肺腺癌发生、发展机制的了解。
虽然本研究结果有助于增进对CDC20与肺腺癌关系的理解,但仍存在一定局限性。首先,对CDC20在肺腺癌中的作用机制的研究是基于生物信息学分析,缺乏基础实验验证。其次,本研究设计有一定局限性,缺少阻断-恢复实验论证,以及动物实验的佐证。未来仍需大规模、全面的研究进一步证实CDC20作为预后生物标志物和治疗靶点的临床疗效。
综上所述,CDC20在肺腺癌中高表达,沉默CDC20表达可抑制肺腺癌细胞增殖、迁移和侵袭,CDC20可能是一种新的肺腺癌预后标志物,进一步的机制研究可为肺腺癌的早期诊断提供新的靶点。
利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:
周雪芹:部分实验操作,撰写文章;
栾艳超:实验设计,修改文章;
赵莉:生物信息学分析;
戎超超:数据搜集、统计及分析;
杨娜:部分实验操作。
[参考文献]
[1] SIEGEL R L, MILLER K D, JEMAL A. Cancer statistics, 2016[J]. CA Cancer J Clin, 2016, 66(1): 7-30.
[2] CASCONE T, FRADETTE J, PRADHAN M, et al. Tumor immunology and immunotherapy of non-small cell lung cancer[J]. Cold Spring Harb Perspect Med, 2022, 12(5): a037895.
[3] CHEN P X, LIU Y H, WEN Y K, et al. Non-small cell lung cancer in China[J]. Cancer Commun, 2022, 42(10): 937-970.
[4] SINGH S S, DAHAL A, SHRESTHA L, et al. Genotype driven therapy for non-small cell lung cancer: resistance, pan inhibitors and immunotherapy[J]. Curr Med Chem, 2020, 27(32): 5274-5316.
[5] CHENG S J, CASTILLO V, SLIVA D. CDC20 associated with cancer metastasis and novel mushroom-derived CDC20 inhibitors with antimetastatic activity[J]. Int J Oncol, 2019, 54(6): 2250-2256.
[6]CHI J J, LI H C, ZHOU Z, et al. A novel strategy to block mitotic progression for targeted therapy[J]. EBioMedicine, 2019, 49: 40-54.
[7]WANG J J, ZHOU F G, LI Y, et al. Cdc20 overexpression is involved in temozolomide-resistant glioma cells with epithelial-mesenchymal transition[J]. Cell Cycle, 2017, 16(24): 2355-2365.
[8]MA X, ZHOU L, ZHENG S S. Transcriptome analysis revealed key prognostic genes and microRNAs in hepatocellular carcinoma[J]. PeerJ, 2020, 8: e8930.
[9]LOVE M I, HUBER W, ANDERS S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2[J]. Genome Biol, 2014, 15(12): 550.
[10]LIU J F, LICHTENBERG T, HOADLEY K A, et al. An integrated TCGA pan-cancer clinical data resource to drive high-quality survival outcome analytics[J]. Cell, 2018, 173(2): 400-416.e11.
[11]YU G C, WANG L G, HAN Y Y, et al. clusterProfiler: an R package for comparing biological themes among gene clusters[J]. OMICS, 2012, 16(5): 284-287.
[12]GIOPANOU I, KANELLAKIS N I, GIANNOU A D, et al. Osteopontin drives KRAS-mutant lung adenocarcinoma[J]. Carcinogenesis, 2020, 41(8): 1134-1144.
[13]贾利晴, 葛小路, 姜 琳, 等. LncRNA PKD2-2-3对肺腺癌细胞增殖、克隆形成、迁移及侵袭能力的影响[J]. 中国癌症杂志, 2023, 33(8): 717-725.
JIA L Q, GE X L, JIANG L, et al. The impact of lncRNA PKD2-2-3 on the proliferation, colony formation, migration, and invasion abilities of lung adenocarcinoma cells[J]. China Oncol, 2023, 33(8): 717-725.
[14]WU Q W, YU L L, LIN X Q, et al. Combination of serum miRNAs with serum exosomal miRNAs in early diagnosis for non-small-cell lung cancer[J]. Cancer Manag Res, 2020, 12: 485-495.
[15]刘晓丽, 柴文君, 孙 磊, 等. 肺腺癌中可变剪接调控因子KHSRP调控的差异剪接基因分析[J]. 中国癌症杂志, 2023, 33(07): 637-645.
LIU X L, CHAI W J, SUN L, et al. Analysis of differentially spliced genes regulated by the alternative splicing factor KHSRP in lung adenocarcinoma[J]. China Oncol, 2023, 33(7): 637-645.
[16]CUADRADO M, GARZÓN J, MORENO S, et al. Efficient terminal erythroid differentiation requires the APC/C cofactor Cdh1 to limit replicative stress in erythroblasts[J]. Sci Rep, 2022, 12(1):10489.
[17]KIM J H, PATEL R. Mad2B forms a complex with Cdc20, Cdc27, Rev3 and Rev1 in response to cisplatin-induced DNA damage[J]. Korean J Physiol Pharmacol, 2023, 27(5): 427-436.
[18]ZHANG Y, XUE Y B, LI H, et al. Inhibition of cell survival by curcumin is associated with downregulation of cell division cycle 20 (Cdc20) in pancreatic cancer cells[J]. Nutrients, 2017, 9(2): 109.
[19]PARMAR M B, K C R B, LÖBENBERG R, et al. Additive polyplexes to undertake siRNA therapy against CDC20 and survivin in breast cancer cells[J]. Biomacromolecules, 2018, 19(11): 4193-4206.
[20]LI K, MAO Y H, LU L, et al. Silencing of CDC20 suppresses metastatic castration-resistant prostate cancer growth and enhances chemosensitivity to docetaxel[J]. Int J Oncol, 2016, 49(4): 1679-1685.
[21]LI J X, WANG Y C, WANG X, et al. CDK1 and CDC20 overexpression in patients with colorectal cancer are associated with poor prognosis: evidence from integrated bioinformatics analysis[J]. World J Surg Oncol, 2020, 18(1): 50.
[22]YANG G, WANG G, XIONG Y F, et al. CDC20 promotes the progression of hepatocellular carcinoma by regulating epithelial-mesenchymal transition[J]. Mol Med Rep, 2021, 24(1): 483.
[23]ZHANG Y Q, LI J Y, YI K K, et al. Elevated signature of a gene module coexpressed with CDC20 marks genomic instability in glioma[J]. Proc Natl Acad Sci U S A, 2019, 116(14): 6975-6984.
[24]WANG L X, ZHANG J F, WAN L X, et al. Targeting CDC20 as a novel cancer therapeutic strategy[J]. Pharmacol Ther, 2015, 151: 141-151.
[25]WANG L X, YANG C L, CHU M, et al. Cdc20 induces the radioresistance of bladder cancer cells by targeting FoxO1 degradation[J]. Cancer Lett, 2021, 500: 172-181.
[26]CHEN Z H, JING Y J, YU J B, et al. ESRP1 induces cervical cancer cell G1-phase arrest via regulating cyclin A2 mRNA stability[J]. Int J Mol Sci, 2019, 20(15): 3705.
[27]AMADOR V, GE S, SANTAMARÍA P G, et al. APC/C (CDC20) controls the ubiquitin-mediated degradation of p21 in prometaphase[J]. Mol Cell, 2007, 27(3): 462-473.
[28]ZHENG Y C, SHI Y, YU S, et al. GTSE1, CDC20, PCNA, and MCM6 synergistically affect regulations in cell cycle and indicate poor prognosis in liver cancer[J]. Anal Cell Pathol, 2019, 2019: 1038069.
[29]KAPANIDOU M, CURTIS N L, BOLANOS-GARCIA V M. CDC20: at the crossroads between chromosome segregation and mitotic exit[J]. Trends Biochem Sci, 2017, 42(3): 193-205.
[30]CHU Z W, ZHANG X Y, LI Q Y, et al. CDC20 contributes to the development of human cutaneous squamous cell carcinoma through the Wnt/β-catenin signaling pathway[J]. Int J Oncol, 2019, 54(5): 1534-1544.
[31]LI J, KARKI A, HODGES K B, et al. Cotargeting polo-like kinase 1 and the Wnt/β-catenin signaling pathway in castration-resistant prostate cancer[J]. Mol Cell Biol, 2015, 35(24): 4185-4198.
[32]KONG J J, WANG X T, WANG J L, et al. Silencing of RAB42 down-regulated PD-L1 expression to inhibit the immune escape of hepatocellular carcinoma cells through inhibiting the E2F signaling pathway[J]. Cell Signal, 2023, 108: 110692.
[33]ZHANG Y, LYU L, WANG W, et al. High expression of E2F transcription factors 7: an independent predictor of poor prognosis in patients with lung adenocarcinoma[J]. Medicine, 2022, 101(33): e29253.
作者:中国癌症杂志
版权声明:
本网站所有注明“来源:梅斯医学”或“来源:MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明“来源:梅斯医学”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言







#肺腺癌# #细胞增殖# #细胞周期# #细胞运动# #细胞分裂周期蛋白20#
80